Top ▲
GtoPdb is requesting financial support from commercial users. Please see our sustainability page for more information.

Target not currently curated in GtoImmuPdb
Target id: 521
Nomenclature: K2P10.1
Abbreviated Name: TREK2
Gene and Protein Information  |
|||||||
| Species | TM | P Loops | AA | Chromosomal Location | Gene Symbol | Gene Name | Reference |
| Human | 4 | 2 | 538 | 14q31.3 | KCNK10 | potassium two pore domain channel subfamily K member 10 | 1 |
| Mouse | 4 | 2 | 535 | 12 E | Kcnk10 | potassium channel, subfamily K, member 10 | |
| Rat | 4 | 2 | 538 | 6q32 | Kcnk10 | potassium two pore domain channel subfamily K member 10 | |
| Previous and Unofficial Names |
| TREK-2 | potassium channel, two pore domain subfamily K, member 10 | potassium channel |
| Database Links |
|
| Alphafold | P57789 (Hs), Q9JIS4 (Rn) |
| ChEMBL Target | CHEMBL2331041 (Hs), CHEMBL3308951 (Rn) |
| Ensembl Gene | ENSG00000100433 (Hs), ENSMUSG00000033854 (Mm), ENSRNOG00000003813 (Rn) |
| Entrez Gene | 54207 (Hs), 72258 (Mm), 65272 (Rn) |
| Human Protein Atlas | ENSG00000100433 (Hs) |
| KEGG Gene | hsa:54207 (Hs), mmu:72258 (Mm), rno:65272 (Rn) |
| OMIM | 605873 (Hs) |
| Pharos | P57789 (Hs) |
| RefSeq Nucleotide | NM_021161 (Hs), NM_029911 (Mm), NM_023096 (Rn) |
| RefSeq Protein | NP_066984 (Hs), NP_084187 (Mm), NP_075584 (Rn) |
| UniProtKB | P57789 (Hs), Q9JIS4 (Rn) |
| Wikipedia | KCNK10 (Hs) |
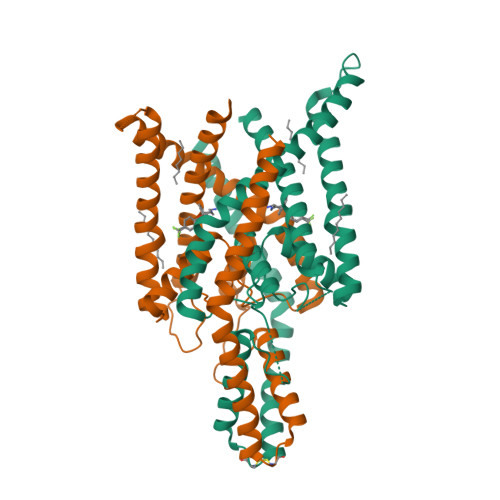
| Selected 3D Structures |
|||||||||||||
|
|
||||||||||||
| Associated Proteins |
||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||
| Associated Protein Comments | ||||||||||||||||||||||||
| Heteromultimers shown to form in vivo: K2P10 has been reported to form a heterodimer with K2P1 in heterologous expression systems, however the in vivo formation of this heterodimer and its function remain unknown [6]. Protein-protein interactions: K2P10 has been reported to respond to numerous proteins, including: AKAP150: The A-kinase-anchoring protein AKAP150 binds to the C-terminus of K2P10, increasing the kinetic of channel inactivation by Gs coupled receptors and decreasing inhibition by Gq coupled receptors [14]. Mtap2: The microtubule associated protein 2 Mtap2 binds to the C-terminus of K2P10 to increase channel density at the plasma membrane [13]. PLD2: PLD2, a phosphatidic acid (PA) producing enzyme, binds to the C-terminus of K2P10 to potentiate channel activity in a PA-dependent manner [2]. |
||||||||||||||||||||||||
| Functional Characteristics |
|
| Background current | |
Download all structure-activity data for this target as a CSV file 
| Activators | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Key to terms and symbols | View all chemical structures | Click column headers to sort | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Activator Comments | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A concentration of 0.5 mM halothane maximally activates the channel by 2.6-fold [9]. The channel is also activated by the neuroprotective drug riluzole. Channel activity is stimulated by 8.4 times by arachidonic acid, and 5.1 times by lysophosphatidylinositol [9]. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Inhibitors | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Key to terms and symbols | View all chemical structures | Click column headers to sort | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Channel Blockers | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Key to terms and symbols | View all chemical structures | Click column headers to sort | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tissue Distribution
|
||||||||
|
| Biologically Significant Variant Comments |
| Three alternate splice forms of K2P10 have been identified [4,9]. To date, the pharmacological and physiological significance of the variant isoforms remains unknown. |
| General Comments |
| ‘Activation’ and ‘deactivation’ with voltage steps appear to be instantaneous. Splice variants have been identified in human and rat. The rat variant is reported to have a conductance of 68pS and to be expressed in brain. K2P10 like currents are observed in cerebellar granular neurons, magnocellular neurosecretory cells of rat supraoptic nucleus [5,8] and rat cortical astrocytes [7]. |
1. Bang H, Kim Y, Kim D. (2000) TREK-2, a new member of the mechanosensitive tandem-pore K+ channel family. J Biol Chem, 275 (23): 17412-9. [PMID:10747911]
2. Comoglio Y, Levitz J, Kienzler MA, Lesage F, Isacoff EY, Sandoz G. (2014) Phospholipase D2 specifically regulates TREK potassium channels via direct interaction and local production of phosphatidic acid. Proc Natl Acad Sci USA, 111 (37): 13547-52. [PMID:25197053]
3. Dong YY, Pike AC, Mackenzie A, McClenaghan C, Aryal P, Dong L, Quigley A, Grieben M, Goubin S, Mukhopadhyay S et al.. (2015) K2P channel gating mechanisms revealed by structures of TREK-2 and a complex with Prozac. Science, 347 (6227): 1256-9. [PMID:25766236]
4. Gu W, Schlichthörl G, Hirsch JR, Engels H, Karschin C, Karschin A, Derst C, Steinlein OK, Daut J. (2002) Expression pattern and functional characteristics of two novel splice variants of the two-pore-domain potassium channel TREK-2. J Physiol (Lond.), 539 (Pt 3): 657-68. [PMID:11897838]
5. Han J, Truell J, Gnatenco C, Kim D. (2002) Characterization of four types of background potassium channels in rat cerebellar granule neurons. J Physiol (Lond.), 542 (Pt 2): 431-44. [PMID:12122143]
6. Hwang EM, Kim E, Yarishkin O, Woo DH, Han KS, Park N, Bae Y, Woo J, Kim D, Park M et al.. (2014) A disulphide-linked heterodimer of TWIK-1 and TREK-1 mediates passive conductance in astrocytes. Nat Commun, 5: 3227. [PMID:24496152]
7. Kang D, Choe C, Kim D. (2004) Functional expression of TREK-2 in insulin-secreting MIN6 cells. Biochem Biophys Res Commun, 323 (1): 323-31. [PMID:15351740]
8. Kang D, Choe C, Kim D. (2005) Thermosensitivity of the two-pore domain K+ channels TREK-2 and TRAAK. J Physiol (Lond.), 564 (Pt 1): 103-16. [PMID:15677687]
9. Lesage F, Terrenoire C, Romey G, Lazdunski M. (2000) Human TREK2, a 2P domain mechano-sensitive K+ channel with multiple regulations by polyunsaturated fatty acids, lysophospholipids, and Gs, Gi, and Gq protein-coupled receptors. J Biol Chem, 275 (37): 28398-405. [PMID:10880510]
10. Loucif AJC, Saintot PP, Liu J, Antonio BM, Zellmer SG, Yoger K, Veale EL, Wilbrey A, Omoto K, Cao L et al.. (2018) GI-530159, a novel, selective, mechanosensitive two-pore-domain potassium (K2P ) channel opener, reduces rat dorsal root ganglion neuron excitability. Br J Pharmacol, 175 (12): 2272-2283. [PMID:29150838]
11. Pope L, Arrigoni C, Lou H, Bryant C, Gallardo-Godoy A, Renslo AR, Minor Jr DL. (2018) Protein and Chemical Determinants of BL-1249 Action and Selectivity for K2P Channels. ACS Chem Neurosci, 9 (12): 3153-3165. [PMID:30089357]
12. Rödström KEJ, Cloake A, Sörmann J, Baronina A, Smith KHM, Pike ACW, Ang J, Proks P, Schewe M, Holland-Kaye I et al.. (2024) Extracellular modulation of TREK-2 activity with nanobodies provides insight into the mechanisms of K2P channel regulation. Nat Commun, 15 (1): 4173. [PMID:38755204]
13. Sandoz G, Tardy MP, Thümmler S, Feliciangeli S, Lazdunski M, Lesage F. (2008) Mtap2 is a constituent of the protein network that regulates twik-related K+ channel expression and trafficking. J Neurosci, 28 (34): 8545-52. [PMID:18716213]
14. Sandoz G, Thümmler S, Duprat F, Feliciangeli S, Vinh J, Escoubas P, Guy N, Lazdunski M, Lesage F. (2006) AKAP150, a switch to convert mechano-, pH- and arachidonic acid-sensitive TREK K(+) channels into open leak channels. EMBO J, 25 (24): 5864-72. [PMID:17110924]
15. Tanaka M, Mori T, Hashimoto G, Mitsui K, Kishi A, Childress ES, Bollinger SR, Chopko TC, Bridges TM, Stafford DG et al.. (2025) Discovery of ONO-TR-772 (VU6018042): A Highly Selective and CNS Penetrant TREK Inhibitor in Vivo Tool Compound. ACS Med Chem Lett, 16 (5): 896-901. [PMID:40365404]
16. Yashiro K, Iwaki Y, Urata H, Kokubo M, Mori T, Sekioka Y, Isami K, Kato J, Wieting J, McGowan KM et al.. (2025) Discovery of ONO-2920632 (VU6011887): A Highly Selective and CNS Penetrant TREK-2 (TWIK-Related K+ Channel 2) Preferring Activator In Vivo Tool Compound. ACS Chem Neurosci, 16 (5): 960-967. [PMID:39981749]